Эмбриогенез кукурузы in vivo
Образование женского гаметофита у растений, в том числе у кукурузы, происходит в пестике, а именно в завязи. В завязи формируется гнездо, в котором развивается семяпочки, в середине которой дифференцируется зародышевый мешок с яйцеклеткой. Пестик закладывается как и другие органы цветка в виде недифференцированного бугорка. Пестик может состоять из одного или нескольких плодолистиков, сросшихся, и в зрелом состоянии имеет завязь, столбик и рыльце. Для кукурузы свойственно образование пестика с одного плодолистиков. Завязь может быть одно-, двух—, трех—и многогнездова. Для кукурузы присуща одногнездова завязь, в которой развивается одна семяпочка [Чеботарь, 1972; Батигина, 1987;Ляпустина 2009]. Развитие и строение сформированного зародышевого мешка большинства покрытосеменных (почти 80% всех эмбриологические—исследованных), в том числе злаков, кукуруза не исключение, соответствует Polygonum—типу по классификации И.Д. Романова (1945), характеризующийся наличием моноспоричного зародышевого мешка, который развивается из нижней клетки тетрады макроспор, имеет восемь ядер и образуется путем пяти делений. И яйцеклетка, и зигота кукурузы имеют четко обозначенную полярность, ядро в них располагается в апикальной зоне клетки, а вакуоль в базальнной. При этом обе инициали имеют грушевидную форму. Согласно многочисленным работам по раннему эмбриогенезу у злаков первые деления в зиготе злаков сопровождаются заложением серии наклонных перегородок, которые приводят к перемещению апекса зиготы из терминального в латеральное положение [Чеботарь, 1972; Батыгина, 1968; Коробова, 1962; Сатарова, 2000; Ляпустина, 2009]. Определяющую роль именно в такой организации яйцеклетки, зиготы и молодого проэмбрио играет их расположение внутри зародышевого мешка и cемязачатка, который отличается анакампилотропностью и дорсовентральностью. Такое строение семязачатка создает градиент поступления питательных веществ через плацентохалазу и приводит к возникновению полярности всех структур семязачатка и элементов зародышевого мешка, что в конечном итоге отражается и на возникновении полярности зародыша с первых этапов его формирования [Сатарова, 2000; Ляпустина, 2009].
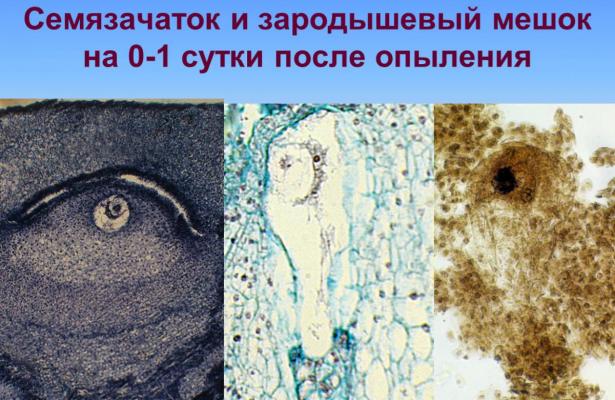
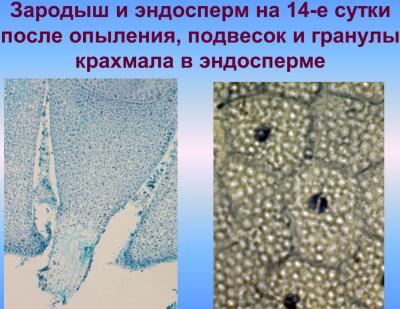
Первичные ядра эндосперма заполняют микропилярную область зародышевого мешка, окружая проембрио, без образования клеточных перегородок. Лишь позже ядра эндосперма займут пристенное положение в зародышевом мешке и еще позже начнет происходить процесс образования клеточных перегородок. Процесс перехода ядерного эндосперма в клеточный, независимо от типа опыления и погодных условий, начинается в микропилярной части зародышевого мешка, то есть непосредственно вокруг проембрио. Ядра эндосперма особенно полиморфные и на раннем этапе содержат по три ядрышка, что связано с их триплоидные природой. Как ядерный так и клеточный эндосперм делятся митотически, выполняя трофическую функцию [Чеботарь, 1972; Ляпустина, 2009].
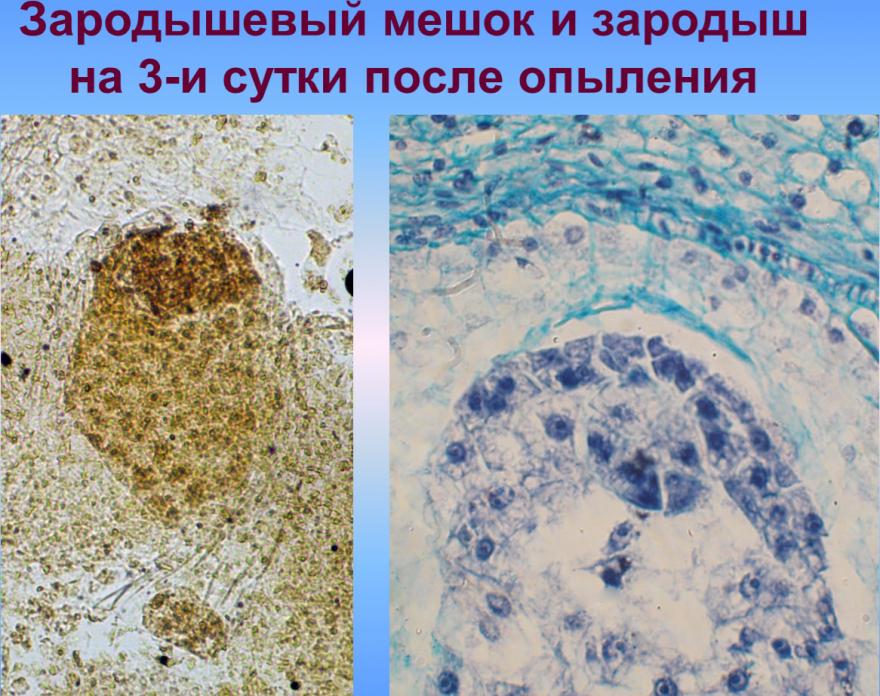
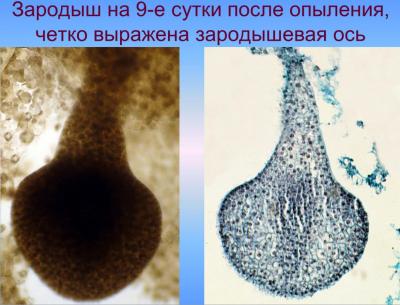
С верхней клетки (апикальной) двухклеточного зародыша (после первого деления) в будущем возникают основные части зародыша, а из нижней (базальной) — подвесок (суспензор). В результате последовательных делений клеток, образуется многоклеточный предзародок с радиальной симметрией, округлой или яйцевидной формы, подобным как для однодольных так и для двудольных. Далее развитие зародыша идет по разным путям для этих двух классов растений. В однодольных, к которым и относится культура кукурузы, образуется только одна конечная семядоля и точка роста стебля закладывается сбоку, благодаря чему зародыш имеет несимметричную форму или одностороннюю симметрию. В двудольных образуются две боковые семядоли, точка роста закладывается в центре между семядолями, зародыш оказывается достаточно симметричным (двусторонняя симметрия). Среди однодольных и двудольных типы развития и строения зародыша варьируют, отличаясь при этом строением подвеска, способом образования перегородок в самом зародыше, форме и величине зародыша и отдельных его частей, размещением зародыша в семенах [Поддубная-Арнольди, 1964; Поддубная-Арнольди, 1964; Тюкавин, 2007]. Развитие полового зародыша в злаков (пшеница, кукуруза) осуществляется в соответствии с Graminad-типа эмбриогенеза [Батыгина, 1997; Круглова, 2005], характеризующееся специфическим наклонным положения первых клеточных перегородок в зародыше, которые возникают в результате асимметричного неравного деления зиготы и обусловливающим дорсовентральнисть его строения с первых этапов развития.
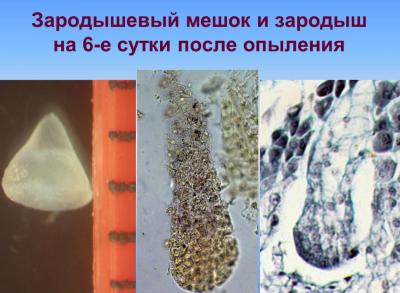
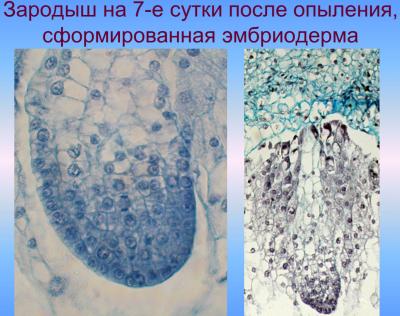
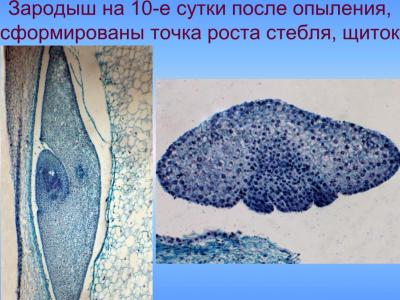
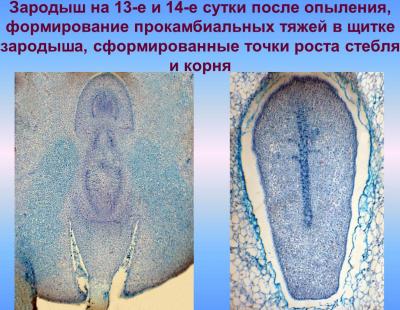
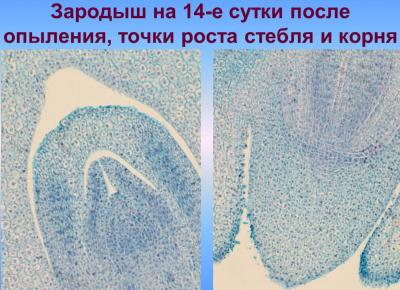
В ходе этой фазы органгогенеза формирования зародыша начинается разрастание его апикально-латеральной области со стороны плаценты-халазы вследствие активных клеточных делений, приводящих к образованию семядоли, или щитка; одновременно с этим процессом происходит закладка эпидермы, которое осуществляется в базипетальном направлении. На другом конце формируется точка роста побега. В процессе органогенеза в зародыше происходят сложные преобразования, в результате которых щиток смещается с латерального положения в терминальное, а точка роста побега — из терминального в латеральное. Следующая стадия — увеличение числа митозов в зоне образования точки роста — апекса побега, а также обособление колеоптиля, возникающий как произрастания основания щитка. Закладка колеоптиле совпадает по времени с дифференциацией прокамбиального тяжа в зародыше и образованием инициалов меристемы главного зародышевого корня. Корень закладывается эндогенно, в базальной части зародыша — в области окончания прокамбиального тяжа, согласно закрытого типа. При этом характерно образование колеоризы, которая возникает как единое образование с чехликом корня. В ходе дальнейших стадий развития колеоптиле постепенно приобретает вид конусообразной трубки с отверстием на вершине; наблюдается закладывание эпибласта со стороны, противоположной щитке, посредством чешуевидными вырасту и зачатка первого листа на апекс ростка.
На поздних стадиях эмбриогенеза наблюдается формирование в апикальной части щитка особого выроста — лигулы (брюшной чешуи), а на его поверхности в области контакта с эндоспермом становится отчетливо дифференцированным особый, эпителиальный, слой, выполняющий функцию абсорбции питательных веществ (гаусториальна функция). На апексе ростка происходит последовательное закладывание зачатков дальнейшего листьев, а также зачатков нескольких (3-5) адвентивных корней. Начинается обособление колеоризы, сопровождающееся изменениями в структуре клеток суспензорий (увеличение их размеров, сильная вакуолизация, деструкция клеток в базальной части суспензорий). Эмбриогенез завершается примерно на 29-30 сутки после опыления. Некоторые различия могут оказываются в терминах закладка органов и времени протекания эмбриогенеза у разных видов и сортов растений [Круглова, 2005; Ляпустина 2009].
Автор статьи и фото в статье: Ляпустина Е.В.
Читайте Эмбриогенез кукурузы in vitro