Особенности процессов дедифференциации и каллусообразования в культуре органов и тканей Clematis vitalba L.
В литературе описывается множество вариантов путей прохождения процессов первичного каллусообразования и морфологическое разнообразие каллусных культур [43, 93, 173].
На рисунках 4.1-4.4 продемонстрированы морфологические изменения экспланта, происходящие в процессе каллусообразования. Так, у изучаемых типов эксплантов Clematis vitalba L. при введении в культуру in vitro первые признаки дедифференциации и каллусообразования были отмечены на 14-28 сутки культивирования и были приурочены к месту поранения экспланта. Процесс образования каллуса начинался раньше у вводимых в условия in vitro высечек физиологически зрелых листьев (иногда на 10-12 сутки, в зависимости от условий культивирования, например, на модифицированных средах В5), затем – у высечек молодых листьев (14-20 сут) и уже в последнюю очередь – у вегетативных почек (20-28 сут).
На эксплантах физиологически зрелых листьев образование каллуса происходило в местах среза экспланта, в частности в местах рассечения листовых жилок. В данном случае процесс каллусообразования был приурочен к месту среза (рис. 4.1, а).
Каллусогенез в культуре вегетативных почек происходил в результате дедифференциации всех тканей экспланта (рис. 4.1, б, в), в ряде случаев образование каллуса не затрагивало физиологически более зрелые ткани экспланта, а формирование первичного каллуса происходило путем разрастания базальной его части и было приурочено к месту среза (рис. 4.1 г).
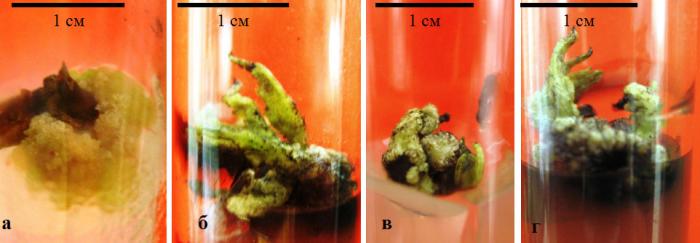
Рис. 4.1. Каллусообразование в культуре эксплантов вегетативных органов Clematis vitalba L.: а – высечек из физиологически зрелых листьев; б – высечек из молодых листьев; в, г – развитие каллуса в культуре апикальной части побега
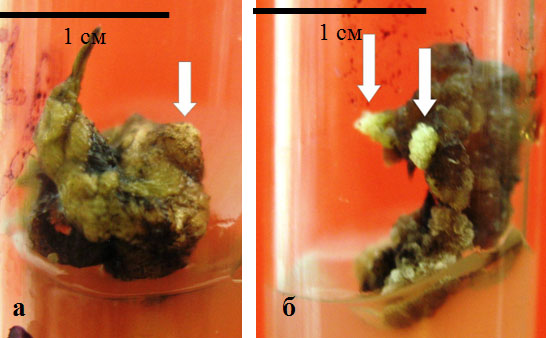
Рис. 4.2. Меристематически активные зоны на поверхности экспланта апикальной части побега (а) и каллуса индуцированного из высечек физиологически зрелых листьев (б) (отмечены стрелками)
В процессе каллусообразования отмечали появление меристематически активных зон, визуально выявляемых в виде бугорков на поверхности экспланта, или каллуса (рис. 4.2, отмечены стрелками).
Образующийся каллус имел разнообразную окраску, которая зависела от состава питательной среды. Так, на средах по прописи LS, с незначительным содержанием регуляторов роста развивался светло-зеленый, плотный, слабооводненный каллус с многочисленными темно-бурыми образованиями (рис. 4.3).
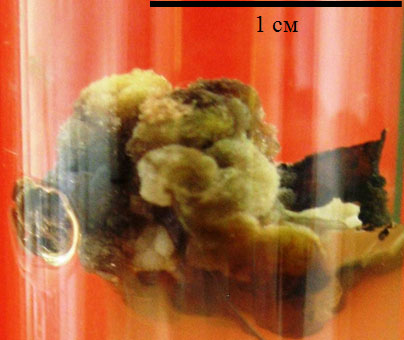
Рис. 4.3. Морфологические особенности каллуса, индуцированного из высечек физиологически зрелых листьев, при культивировании на модифицированной (1,0 мг/л 2,4-Д и 0,5 мг/л БАП) среде LS
На средах по прописи В5, содержащих 2,4-Д и БАП до 3,0 мг/л, развивался плотный, светлый, слабооводненый, по мере достижения стационарной фазы темнеющий каллус с многочисленными светлыми образованиями, за счет которых и происходил его рост (рис. 4.4, а, рис. 4.4, б).
На вариантах сред, содержащих высокие концентрации регуляторов роста (более 3,0 мг/л), происходило образование темного бугристого каллуса, разрастающегося небольшими зонами, не отличающимися по цвету и морфологии от основной массы каллусной ткани (рис. 4.4, в).
При культивировании в темноте на средах по прописи В5 каллусные культуры приобретали более рыхлую консистенцию, в начале имели светлую, а затем – слегка буроватую окраску (рис. 4.5, б). На свету отмечена более низкая скорость роста каллуса, который приобретал светло-зеленую окраску, с постепенным потемнением ткани при достижении стационарной фазы (рис 4.5, а).

Рис. 4.4. Морфология каллусной ткани, индуцированной из высечек листьев, при культивировании напитательных средах по прописи В5: а – в начале культивирования; б – в середине цикла выращивания (концентрация регуляторов роста до 3,0 мг/л); в – в середине цикла выращивания (концентрация регуляторов роста в среде свыше 3,0 мг/л)

Рис. 4.5. Морфологичеcкие различия каллусов Clematis vitalba L., индуцированных из высечек физиологически зрелых листьев, при культивировании на питательной среде В5 на свету (а) и в темноте (б)
Каллус, сформировавшийся из эксплантов разного типа, на средах по прописи МС на свету имел светлую, светло-зеленую окраску, плотную консистенцию и невысокую скорость роста (рис. 4.6, а). В темноте цвет каллуса менялся от светло- до темно-бурого (рис. 4.6, б). При выращивании на данной среде каллус характеризовался более низкими показателями прироста биомассы, чем при культивировании на среде В5. Образовавшийся на свету каллус отличался относительной гомогенностью структуры, в темноте же была отмечена слабая степень его гетерогенности.

Рис. 4.6. Каллус Clematis vitalba L., индуцированый из высечек физиологически зрелых листьев, при культивировании на питательной среде МС на свету (а) и каллус, индуцированый из апикальной части побега, при культивировании на среде МС в темноте (б)
В результате пассирования на свежеприготовленные питательные среды каллусные культуры сохраняли невысокую интенсивность роста и морфологическую неоднородность. Вне зависимости от условий культивирования (в темноте или на свету) и состава питательной среды (МС, В5 или LS) по достижении стационарной фазы каллусные культуры приобретали более рыхлую консистенцию и буроватую окраску.
Наблюдения за ростом каллусных культур показали, что наряду с нарастанием самого каллуса проявлялись и признаки вторичной дифференциации: примерно на 90 сутки на поверхности каллусов в вариантах питательных сред по прописи МС и В5 появлялись зачатки корней, которые при дальнейшем культивировании развивались в нормальные корни (рис. 4.7). На средах по прописи LSморфогенез в каллусных культурах не был выявлен.
Таким образом, наши данные свидетельствуют о том, что в культуре вегетативных эксплантов Clematis vitalba L. каллусообразование происходит путем дедифференциации эксплатнов по месту среза или дедифференциации всех тканей экспланта. Образующийся в результате дедифференциации экспланта каллус имеет разнообразную морфологию и характеризуется низкой скоростью роста.

Рис. 4.7. Ризогенез в каллусных культурах, индуцированых из высечек физиологически зрелых листьев Clematis vitalba L.: а – на свету; б – в темноте (95 сут культивирования); в – в темноте (150 сут культивирования)
Визуальные наблюдения за процессами каллусообразования на средах оптимизированных по матрице эксперимента показало сходство их протекания с вышеописанными процессами. Так же, как и в описанных ранее процессах, морфология образующегося каллуса в большой степени зависела от состава питательной среды и условий культивирования.
Как показали наши исследования, при культивировании в условиях освещения и в термостате (в темноте) наиболее интенсивно прирост биомассы происходил на оптимизированных питательных средах № 1, 2, 3, 11, 12, 13 и 23.
Цикл выращивания первичных каллусных культур на средах по оптимизированным прописям составлял от 75-ти до 90-та суток. Однако, при проведении субкультивирований нами выявлена незначительная интенсификация скорости роста каллуса, начиная с 4 пассажа, цикл выращивания каллусов сократился до сократился до 60-ти–75-ти суток.
Наши исследования показали, что при культивировании на свету образующийся каллус характеризовался компактной структурой, слабой степенью оводненности и невысокой интенсивностью роста (рис. 4.8).
При культивировании на средах № 1 и № 2 (рис. 4.8, а, б), формирующийся каллус имел плотную консистенцию и приобретал темную (от светло-коричневого до темно-бурого, почти черного) окраску со светлоокрашенными «ростовыми» зонами.
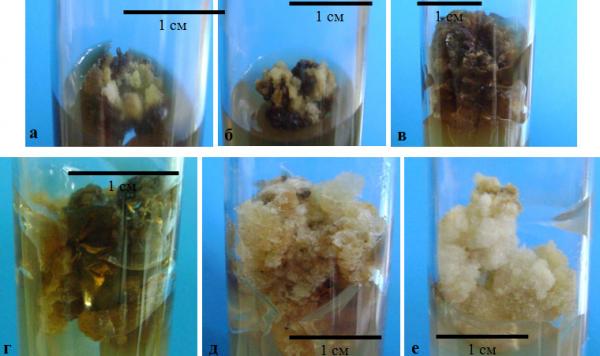
Рис. 4.8. Морфологическое разнообразие каллусных культур, индуцированых из высечек физиологически зрелых листьев, при культивировании их на различных вариантах питательных сред в условиях освещенности: а – среда № 1, б – среда № 2, в – среда № 3, г – среда № 11, д, е – среда №12
При культивировании на средах № 3 и № 11 (рис. 4.8, в, г) каллус характеризовался рыхлой консистенцией, высокой степенью оводненности и скоростью роста и имел по сравнению с каллусами, выращиваемыми на средах № 1 и № 2, более светлую, коричневатую или светло-бурую окраску. При этом на среде № 11 вновь образующийся каллус был способен к разрастанию по поверхности питательной среды и по достижении стационарной фазы опускался по направлению к дну пробирки. Необходимо отметить и тот факт, что на данных вариантах сред по достижении каллусом стационарной фазы роста питательные среды приобрели темную, иногда почти черную окраску за счет выделения каллусными тканями пигментов.
Наиболее интенсивная продукция пигментов была отмечена при культивировании каллуса на среде по прописи № 1, менее интенсивное выделение пигментов наблюдали на средах № 2, № 3 и самое низкое – на среде №11 (рис. 4.8, а-г).
Каллус, формирующийся на среде № 12 (рис. 4.8, д, е), имел более рыхлую консистенцию, высокую скорость роста и отличался светлой (беловатой, иногда желтоватой или светло-серой) окраской. В данном варианте культивирования выделения в питательную среду пигментов не выявлено. Признаков проявления морфогенетической потенции каллусными культурами в случае выращивания их на вариантах питательных сред № 1, 2, 3, 11 и 12 невыявлено.
Морфология полученных первичных и пассируемых каллусных культур при их выращивании на оптимизированных средах в темноте представлена на рисунках 4.9-4.12. Так, каллус, выращиваемый на среде № 1 (рис. 4.9, а, б), сохранял невысокую скорость роста, но отличался от культивируемых на свету более светлой окраской (от светло-коричневой до коричнево-бурой), большим количеством светлоокрашенных «ростовых» зон и более низкой способностью к продуцированию и выделению в среду культивирования пигментов по достижении стационарной фазы роста. Однако каллус сохранял свою плотность, низкий уровень оводненности и довольно низкие ростовые показатели.
При выращивании каллусов на среде № 2 в темноте их характерными особенностями были невысокая скорость роста, окрашивание в серый цвет, низкая оводненность, отсутствие выделения пигментов в питательную среду или же слабая способность к продуцированию пигментов в конце цикла выращивания (рис. 4.9, б).

Рис. 4.9. Морфологическое разнообразие каллусов, индуцированых из высечек физиологически зрелых листьев, при культивировании на различных вариантах питательных сред в темноте: а, б – среда № 1, в – среда № 2
При выращивании каллуса на среде № 3 были выделены три его основные типа:
- темно-бурый, с беловатыми зонами активного роста, не способный или проявляющий низкую способность к выделению в питательную среду пигментных соединений, с высокой степенью оводненности, рыхлый по консистенции и с довольно высокой интенсивностью роста (рис. 4.10, а).
- каллус, имеющий коричневатый (карамельный, иногда светло-бурый) цвет с зонами роста, окрашенными в светло-коричневые или желтоватый цвета, характеризующийся довольно низкой интенсивностью роста, средней плотности, невысоким уровнем оводненности, способный к продуцированию и выделению в питательную среду пигментных соединений (рис. 4.10, б).
- каллус светлый, беловатый, по мере разрастания окрашивающийся в светло-коричневый цвет, с невысокой скоростью роста, рыхлый, с высокой степенью оводненности, не способный к выделению в среду пигментных соединений (рис. 4.10, в).

Рис. 4.10. Морфологическое разнообразие каллусов, индуцированых из высечек физиологически зрелых листьев, при культивировании их на питательной среде № 3 в темноте
При культивировании на среде № 11 в отсутствии освещения (рис. 4.11, а) каллус был светлоокрашенным, желтоватым, плотным, с невысокой скоростью роста, не вызывающим пигментацию питательных сред.
При культивировании эксплантов на среде № 12 (рис. 4.11, б, в) в темноте образовывался каллус рыхлый по консистенции, светлоокрашенный, по мере достижения стационарной фазы роста иногда темнеющий, с довольно высокой интенсивностью роста, высокой степенью оводненности, не продуцирующий пигментов в питательную среду.
Для сред № 13 и 23 при культивировании в темноте отмечен более интенсивный, чем в условиях освещенности рост. Формирующийся при этом каллус обладал следующими морфологическими особенностями: на среде № 13 (рис. 4.12, а) образовывался рыхлый, сильно оводненный, однородный каллус темно-бурого, почти черного цвета, с низкой интенсивностью роста, не выделяющий в питательную среду пигментов либо выделяющий их в очень малых количествах. На среде № 23 (рис. 4.12, б) образовывался каллус с невысокой скоростью роста, рыхлый, средней степени оводненности, беловатой окраски приобретающий серый цвет, по мере старения каллуса иногда отмечали появление карамельного оттенка. Выделения каллусом пигментов в питательную среду не наблюдали.

Рис. 4.11. Морфологическое разнообразие каллусов, индуцированых из высечек физиологически зрелых листьев, при культивировании на различных вариантах питательных сред в темноте: а – среда № 11, б – среда № 12 в середине цикла выращивания (4 нед. культивирования), в – среда № 12 в конце цикла выращивания (8 нед. культивирования)

Рис. 4.12. Морфологическое разнообразие каллусов, индуцированых из высечек физиологически зрелых листьев, при культивировании их на различных вариантах питательных сред в темноте: а – среда № 13, б – среда № 23
Таким образом, проведенные исследования показали, что в культуре изолированных тканей Clematis vitalba L. первые признаки дедифференциации и индукции каллусообразования отмечаются у изучаемых типов эксплантов на 14-28 сутки культивирования, приурочены к месту нанесения травматического воздействия, а морофлогический тип образующегося каллуса определяется трофическими, регуляторными и физическими условиями культивирования.
Ссылки на литературу и рисунки в тексте соответствуют источнику Индуцированный морфогенез in vitro и накопление тритерпеновых гликозидов в каллусных культурах ломоноса виноградолистного (Clematis vitalba L.)
Источник: Сидякин А.И., Индуцированный морфогенез in vitro и накопление тритерпеновых гликозидов в каллусных культурах ломоноса виноградолистного (Clematis vitalba L.): Дис. … канд. биол. наук. Симферополь. 2011. – 217 с.
Материал любезно предоставил Сидякин Андрей Иванович – к.б.н., ас. каф. ботаники и физиологии растений и биотехнологий и м.н.с. Биотехнологического центра НИЧ Таврического национального университета им. В.И. Вернадского, научный консультант ООО КрымБио