Морфолого — культуральные признаки Saccharomyces carlsbergensis
Saccharomyces carlsbergensis — пивные дрожжи низового брожения. Форма клеток лимоноподобная, клетки могут достигать 20-25 мкм.
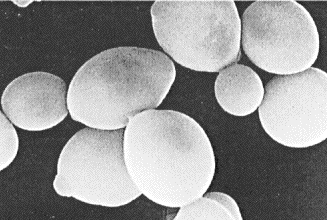
Рисунок 1 — Клетки Saccharomyces carlsbergensis
Клетки суточной культуры на жидком пивном сусле овально-округлой формы, размере 7,0 × 8,4 мкм.

Рисунок 2 — Рост Saccharomyces carlsbergensis на жидкой питательной среде
Вызывают помутнение жидкости. По окончании брожения среда становится более прозрачным, дрожжи оседают на дно, образуя плотный осадок желтовато-белого цвета, отделяется при сильном взбалтывании комками. Пленка на поверхности среды не развивается.
На сусло-агаре — клетки дрожжей овальной формы размером 7-8 × 12-13 мкм.
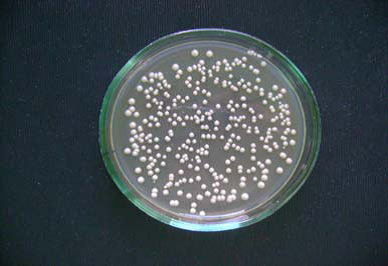
Рисунок 3 — Колонии Saccharomyces carlsbergensis на агаризованной среде
Макроколония на агаре (20 сутки выращивания) округлой формы, блестящая, профиль кратеровидный, поверхность колонии радиально исчерченность с темно-коричневыми концентрическими кругами, край волнистый, структура однородная, консистенция мажущаяся. Штрих на косом сусло-агаре выпуклый, с ровными краями, желтовато-белого цвета, маслянистый.
Внесение триптофана в среду способствует формированию бархатистых колоний Saccharomyces carlsbergensis. Эндоспоры не образуют.
Дрожжевая клетка Saccharomyces carlsbergensis отграничена от внешней среды прочной эластичной стенкой (оболочкой), толщина которой составляет 70 -130 нм. Клеточная стенка содержит гемицеллюлозы (до 60-70% сухой массы), в состав которых входят глюканы, Манана и хитин.
Мананы и глюканы являются основными полимерами клеточной стенки в зависимости от условий культивирования они составляют 60-80% сухой массы стенки. Содержание хитина в клетках аскомицетних дрожжей, почкующихся, незначительный, всего 1-2% от сухой массы клеточной стенки, находится он главным образом в почках.
Много дрожжей, живущих в природных субстратах, особенно бедных доступными питательными веществами, формируют на поверхности клетки капсулу. Раньше думали, что капсула обеспечивает прикрепление клеток, препятствуя их смыванию, а также защищает клетку от воздействия неблагоприятных факторов внешней среды. Более поздние исследования показали, что наличие капсулы обеспечивает клетке утилизацию веществ при очень низкой их концентрации в окружающей среде.
Для капсул Saccharomyces carlsbergensis характерна более четкая ориентация фибрилл и строго параллельное их расположение.
Физиолого-биохимические признаки Saccharomyces carlsbergensis
Дрожжи Saccharomyces carlsbergensis являются факультативными анаэробами, обладают способностью сбраживать сахара при недостатке кислорода в среде и окислять эти сахара при достаточном количестве кислорода.
Скорость утилизации глюкозы в них достигает 100-300 мкмоль-ч, а скорость потребления кислорода — всего 5-50 мкмоль. Интересно отметить, что особенно низкая скорость потребления кислорода обнаружена в штаммов Saccharomyces carlsbergensis.
У Saccharomyces carlsbergensis скорость роста на глюкозных средах низкая. В этих дрожжей глюкоза репрессирует дыхательные ферменты и потому брожения является главным катаболическим путем превращения сахаров, даже в аэробных условиях. У них не наблюдается больших различий в скорости потребления сахара зависимости от присутствия или отсутствия кислорода. Небольшой эффект Пастера может проявляться только в специальных экспериментальных условиях, например в хемостатний культуре в стационарной фазе, или в клетках, сбраживают сахар в отсутствие источника азота. В нормальных условиях роста, при избытке в среде сахара и азотного питания в этих дрожжей пастеривский эффект не проявляется.
Наиболее благоприятное значение рН среды для роста дрожжей находится в пределах 3,5-5,0. Вероятно, эти границы рН совпадают с оптимальным значением рН для активности ферментов, связанных с плазматической мембраной.
Добавление в питательную среду сахара (15%), или соли (выше 1-1,5%) негативно влияет на жизнедеятельность дрожжей Saccharomyces carlsbergensis. Этиловый спирт подавляет их развитие в концентрации 2-5%. При содержании 12-14% спирта, брожение в большинстве случаев прекращается.
При действии глицерина на дрожжи Saccharomyces carlsbergensis снижается водная активность, тем самым снижается разница осмотического давления по обе стороны плазматической мембраны.
Установлено, что при концентрации выше 1 ммоль / л и рН ниже 4,0 сульфит вызывает быстрое уменьшение количества АТФ в клетках дрожжей Saccharomyces carlsbergensis, что приводит к их гибели. Если дрожжи выдерживать с сульфитом менее 1 ч, то снижение АТФ обратимое и клетки не погибают.
Сульфит вызывает ингибирование или инактивацию глицеральдегид-3-фостатдегидрогеназы — фермента, что приводит к накоплению глицеральдегид-3-фосфата и активации АТФ-гидролитических системы, что в свою очередь ведет к уменьшению количества АТФ.
Дрожжевая клетка Saccharomyces carlsbergensis состоит из клеточной стенки, цитоплазмы и органелл. Цитоплазма состоит из воды, различных белков, углеводов, липидов, минеральных веществ, определяющих ее вязкость. Цитоплазма осуществляет ряд важных функций, связанных с ростом клетки, в процессах транспорта. В цитоплазме содержатся структурные компоненты — органеллы, каждая из которых выполняет определенную физиологическую и биохимическую функцию. К органелл относятся ядро, митохондрии, рибосомы, аппарат Гольджи, эндоплазматический ретикулум, лизосомы, вакуоли, микротельца, фаголизосомы, микротрубочки.
Ядро можно наблюдать в живых клетках дрожжей, используя фазово-контрастную и люминесцентные микроскопию, на фиксированных и окрашенных препаратах, а также при электронной микроскопии.
Диаметр ядра составляет 1-2 мкм. Обычно оно имеет сферическую форму, но в тех случаях, когда вакуоль оттесняет ядро к клеточной стенке, оно может деформироваться и становиться серповидным или веретенообразным. У клеток, почкующихся, ядро, как правило, располагается между почкой и вакуоли; сразу же после отделения почки оно перемещается в противоположную сторону материнской клетки. Ядро является главным компонентом клетки, так как именно в нем содержится почти вся генетическая информация клетки.
Митохондрии — высокоспециализированные органеллы. Количество митохондрий в клетке изменяется от 1 до 50 и зависит от условий культивирования и физиологического состояния. Каждая митохондрия отграниченная двойной мембраной. Внешняя мембрана гладкая, внутренняя образует кристы. Размер и количество крист в значительной степени зависит от метаболического состояния клетки. Основная функция митохондрий заключается в выделении энергии путем окисления субстратов и накопления этой энергии в форме АТФ.
Рибосомы — ультрамикроскопические сферические гранулы, размером 20 нм. Для дрожжей характерные 80S-рибосомы, на их долю приходится до трети массы клетки.
Аппарат Гольджи оказывается в клетке Saccharomyces carlsbergensis очень редко. Он служит для секреции продуктов метаболизма клетки.
Эндоплазматическая сеть представляет собой систему мембранных пластин, канальцев, цистерн, окруженная мембраной. Мембрана сетки бывает двух типов: шероховатая и гладкая. Шероховатая эндоплазматическая сеть осуществляет внутриклеточный транспорт. Гладкая эндоплазматическая сеть участвует в обмене углеводов, синтезе липидов. Одна из главных функций эндоплазматической сети в дрожжевой клетке — участие в локальном росте клеточной стенки.
Лизосомы — тельца, окруженные липопротеидной мембраной. Они содержат большой набор ферментов, гидролизуют белки, полисахариды и нуклеиновые кислоты.
Вакуоли являются производными эндоплазматического ретикулума или аппарата Гольджи. Размеры и форма вакуолей — неодинаковы. В цитоплазме молодых клеток дрожжей содержится обычно одна вакуоль, если они имеют сферическую или эллипсоидальной формы. В вакуолях находятся белки, положительно заряженные аминокислоты, липиды, углеводы и некоторые другие вещества. Вакуоли играют важную роль в явлениях осмоса благодаря способности накапливать в большом количестве ионы Na +, К +, Са2 +, Mg2 +, Cl-, SO42-В цитоплазме и вакуолях клеток обнаруживаются скопления гликогена, метахроматина (волютина), липидов, серы, сахаров и кристаллов кислот. Они находятся в клетке в осмотически-инертной форме и обычно не растворимые в воде.
Трегалоза и гликоген содержатся в клетках дрожжей в значительных количествах и является запасными веществами. Трегалоза преобладает у дрожжей, выращенных в аэробных условиях, а гликоген — в анаэробных (содержание его в некоторых случаях может составлять до 40% сухой массы клеток).
В начале брожения гликоген появляется в клетках в виде небольших включений, разбросанных в цитоплазме, которые затем сливаются в большие гранулы. Наличие гликогена легко обнаружить после обработки клеток раствором Люголя. К концу брожения он постепенно исчезает. Когда дрожжи спорулюють, гликоген накапливается в асках и используется аскоспорами во время их прорастания.
По типу питания Saccharomyces carlsbergensis — хемоорганогетеротрофный микроорганизм.
При ДНК-ДНК гибридизации Saccharomyces carlsbergensis обнаруживает родство и с Saccharomyces cerevisiae (57%), и с Saccharomyces bayanus (72%). Ученые полагают, что Saccharomyces carlsbergensis является частичным амфидиплоидом, которое может возникать при естественной гибридизации между S. cerevisiae и S. bayanus.
Размножение Saccharomyces carlsbergensis происходит вегетативным или половым способом. Наиболее благоприятной для спорообразования является температура 25°С. По мнению многих исследователей, лучшем спорообразования способствует щелочная реакция среды. На ацетатном агаре оптимум рН для спорообразования штамма Saccharomyces carlsbergensis составляет 8,4-10,6. На глюкозы агаре хорошо спорообразования пивных дрожжей наблюдается в пределах рН 7,0-8,4, в то же время на агаре из пирувата натрия лучшие результаты получены при рН 4,8-8,4.
Плотность клеточной популяции для спорообразования должна быть не менее 0,5 × 106 кл. / Моль агара, хотя влияние этого показателя зависит от состава среды и условий культивирования. Благоприятно влияет присутствие молекулярного кислорода, в то время как диоксид углерода в высокой концентрации сильно угнетает спорообразования.
Установлено, что ионы К + стимулируют спорообразования Saccharomyces carlsbergensis большей степени, чем Na +. Ионы K + не проявляют токсичности даже в высокой концентрации, a Na + и особенно Са2 + подавляют спорообразования в значительно меньшей концентрации.
По полового размножения в результате мейотического деления диплоидного ядра дрожжей образуются гаплоидные споры — аскоспоры и споридии. Чаще всего в АСКУ содержится 1-8 спор. Аскоспоры бывают 2-х видов спаривание:
- а
- α
Вследствие спаривания гаплоидных клеток но и α образуется нормальная диплоидная клетка а / α. Гаплоидные клетки одного типа также могут случайно спариваться, образуя аномальные диплоидные клетки а / а или α / α, которые размножаются только обычным бесполым путем — почкованием.
На процесс аскообразования влияет ряд факторов. Способность к споруляции зависит от стадии развития вегетативной клетки: только образованные клетки спорулюют медленнее, чем клетки, взятые в середине вегетативного клеточного цикла. Наиболее интенсивно спорулюют клетки, находящиеся в фазе последней трети клеточного цикла. Материнские клетки спорулюют лучше, чем дочерние. Одной из предпосылок для максимальной споруляции есть предварительная адаптация клеток к метаболизму.
У Saccharomyces carlsbergensis вегетативное размножение происходит путем многостороннего почкования, т.е. формирование почек на разных сторонах материнской клетки. Процесс почкования заключается в том, что в клетке появляется небольшой бугорок — почка. С ростом дочерней клетки перепонка, соединяющая его с материнской клеткой, сужается. Примерно через 2 часа молодая клетка отделяется от старой, оставляя на месте своего образования четкий рубец, структура которого зависит от способа вегетативного размножения. В неблагоприятных условиях образуются несколько почек, поскольку молодые клетки не отделяются от старой.
В течение жизни материнская клетка почкуется 25-30 раз. При вегетативном размножении ядро клетки делится путем митоза, т.е. в поколениях клеток сохраняется строго постоянное число хромосом. Различные дрожжи имеют неодинаковое количество хромосом. Считают, что в
Saccharomyces carlsbergensis в гаплоидному состоянии количество хромосом равно 17. В диплоидных клетках их количество удваивается.
Таксономическое положение Saccharomyces carlsbergensis
В 1984 г. была опубликована классификация Н. Дж. Крегер-ван Рой, которая на протяжении почти 16 лет считалась самой полной. Согласно этой классификации Saccharomyces carlsbergensis относятся к отделу Eumycota, подотдела Ascomycotina, класса Hemiascomycetes, порядка Endomycetales, семьи Saccharomycetaceae.
Согласно филогенетической классификации дрожжей К. П. Куртцман, Saccharomyces carlsbergensis относятся к классу Hemiascomycetes, порядка Saccharomycetales, семьи Candidaceae.
Таблица – КласификацияКуртцмана (2000 г.)
| Класс | Порядок | Семейтство | Род |
| Hemiascomycete | Saccharomycetales | Candidaceae |
Brettanomyces,Candida, Saccharomyces |
| Базидиомицетные дрожжи | Sporobolomycetaceae | Sporobolomyces, Rhodotorula | |
| Cryptococcaceae |
Burelia, Cryptococcus,Trichosporon |
||
Материал статьи любезно предоставила Просяник Алина.